
This month’s ”Group of the month” is a bit of shameless self-promotion. I will present an animal group today, where we conduct quite a lot of research on. However, it is nonetheless really interesting and worth to learn more about it. The family Parergodrilidae belongs to the annelids and hence is related to earthworms, bone-eating worms or rag-worms. Its members are rather small with body sizes of about 1 to 3 mm and they are separated into two genera, Parergodrilus and Stygocapitella. Stygocapitella occurs in medium coarse sediments in the damp zone of the upper shore of sandy beaches but occasionally also in sediments with higher water content, while Parergodrilus can be found in the humus-rich upper layer of soil, often within the first three to six centimeters. Hence, this is one of the many examples in animals of a transition from a marine life to a terrestrial one, whereby Stygocapitella still exhibits the transitory stage living in a habitat influenced by both marine and terrestrial environmental factors.
In contrast to many other terrestrialization events, this one did not lead to a strong radiation of species and occupation of different terrestrial habitats. So far, only one species of Parergodrilus is known and it occurs all over the Northern hemisphere, with most records from Europe. It is not certain yet if occurrences outside Europe are due to human-mediated transport and hence the species is invasive outside Europe or if it reflects biased sampling activities. However, it is very likely that the terrestrialization happened a long time ago as the two genera split at least 260 million years ago. Hence, it appears that enough time would have passed to allow for a more prolific radiation and adaptation to terrestrial habitats. Why the radiation is still so limited and restricted is unknown and requires further investigations. On the other hand, the species did also not go extinct, which would indicated that it is very well adapted to its niche and resilient to change. Moreover, if it is truly invasive outside Europe, this would indicate that it has also a good capacity to adapt and thrive in new environments.

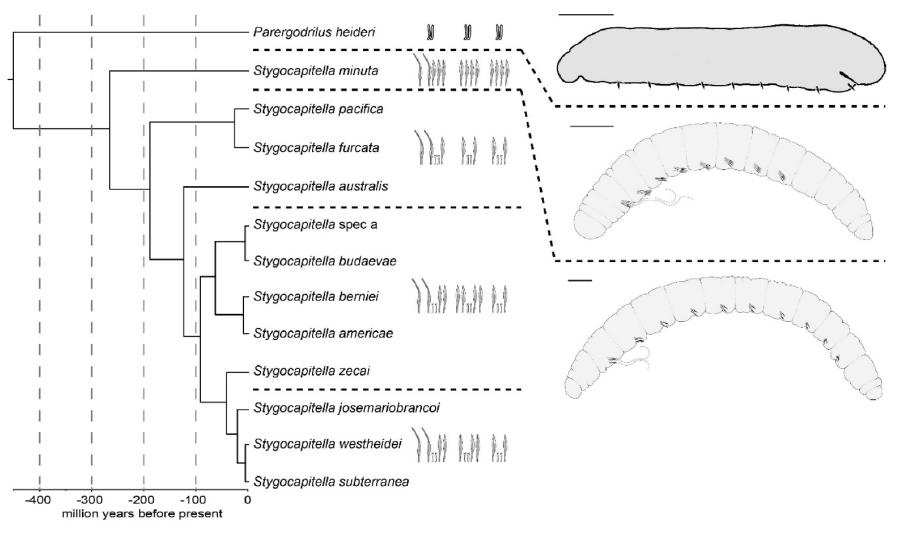
The other genus Stygocapitella is more widely distributed and occurs both on the Northern and Southern hemisphere, but with fewer records in the latter and none in the Tropics and Subtropics. Until recently, only a single species has been described, but increased efforts in both sampling of different localities and integrative taxonomic studies using both morphological and molecular data revealed that at least 12 species occur in this genus. It is a prime example of cryptic species as the 12 species can only be differentiated in four different morphotypes. Species within the morphotypes are absolutely identical and even between three of the four morphotypes the differences are extremely limited with only a one to three chaetae being present or lacking in the first two segments. Interestingly, specimens for morphologically identical species occur in a cup-sized sample from the same beaches (e.g., the beach of Musselburgh harbor) together, but no hybridization happens between them. Hence, while we cannot differentiate the species, the species can recognize each other as separate. The question is how do they do that? Moreover, how can they occupy apparently the same niche without out-competing one another?
The results also showed that Stygocapitella is no longer a cosmopolitan species as assumed for a long time. This always posed a challenging question how this one species could disperse so good as it had no effective dispersal stage. In specific, no pelagic larva, no swimming capacity as an adult, no sperms or eggs released to the open water, also passive dispersal opportunities are very limited given its habitat so high up the beach. This phenomenon is known as the Meiofauna paradox as it is reported more often for such small species. The more restricted distributions after the taxonomic revision seem to reduce this paradox substantially, but still the animals can exchange genetic information over hundreds of kilometers, as for example specimens along the entire US-Atlantic coastline share the same mitochondrial genome sequence. However, one very interesting question remains despite the more restricted distribution. Given the lack of any record from the Tropics and Subtropics, how can the genus be distributed in both the Southern and Northern hemisphere? Given recent molecular data, Stygocapitella species must have crossed the equator at least twice. How did they manage to travel thousands of kilometers through a seemingly hostile environment?
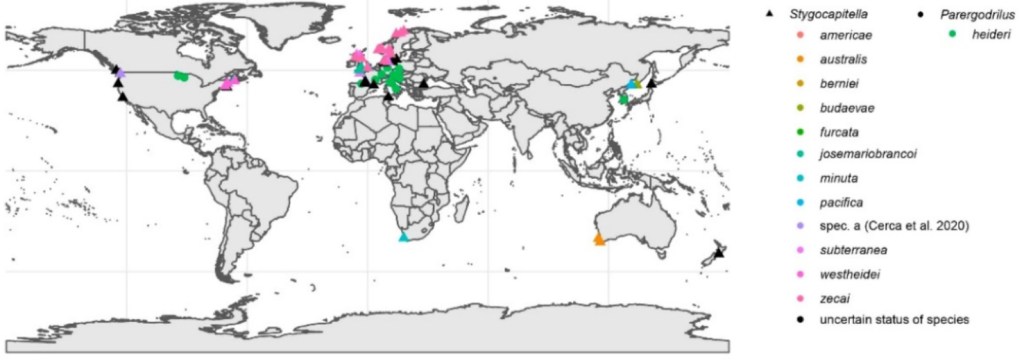
Last, but by really now means least, is another aspects of these fascinating Stygocapitella species. One of the three morphotypes, which are very similar to each other, can be found in three species from Russia, the US-Pacific and Australia and it evolved about 180 million years ago. The second one of the three is found in five species from Russia, the US-Pacific and Europe and has been around for about 80 million years, while the last one found in three species from Europe and the US-Atlantic originated about 20 million years ago. Hence, their morphological evolution is very static and some originated while the dinosaurs were still around. The dinosaurs went extinct, but they are still here. In paleontology, this phenomenon is known as morphological stasis and it is still a challenge in evolution to explain why species do not change for such a long time. Given all the environmental changes and just by chance, some change and differences should occur and when it does not it requires a reason. While there is no shortage of possible explanations, they can often not be thoroughly tested as genetic and detailed ecological data are lacking. The Stygocapitella species with these extreme cases of stasis provide hence a perfect system to approach this problem from a new perspective allowing the inclusion of detailed genetic, morphological and ecological data. Why do no morphological difference accumulate despite tens of millions of generations?
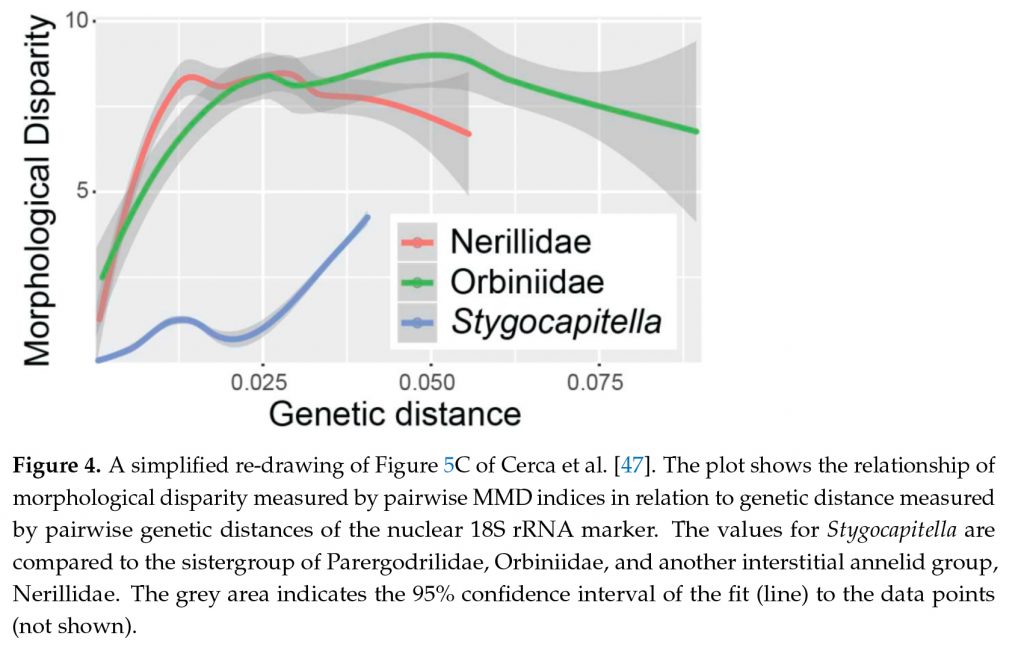
Hence, even though it is a tiny family with few species it poses nonetheless several interesting questions about evolution, species recognition and species distribution. The most captivating one is probably why so little is happening be it after terrestrialization or due to morphological stasis and maybe one is connected with the other. These cases of stable systems are also topic of our newly established research hub STADIS (Stability and Discontinuity) at the NHM.
Further reading:
Struck, T. H. & Cerca, J. (2022). Connecting micro- and macro-evolutionary research – extant cryptic species as systems to understand macro-evolutionary stasis. In: Munro, A. (Editor) Cryptic species. Cambridge University Press, Cambridge.
Meca, M. A., Zhadan, A. & Struck, T. H. (2021). The Early Branching Group of Orbiniida Sensu Struck et al., 2015: Parergodrilidae and Orbiniidae. Diversity 13: 29. (see also our blog entry)
Cerca, J., Rivera-Colón, A. G., Ferreira, M. S., Ravinet, M. Nowak, M. D., Catchen, J. M. & Struck, T. H. (2021). Incomplete lineage sorting and ancient admixture, and speciation without morphological change in ghost-worm cryptic species. PeerJ 9: e10896. (see also our blog entry)
Cerca, J., Meyer, C., Stateczny, D., Siemon, D., Wegbrod, J., Purschke, G., Dimitrov, D. & Struck, T. H. (2020). Deceleration of morphological evolution in a cryptic species complex and its link to paleontological stasis. Evolution 74: 116–131.
Cerca, J., Meyer, C., Purschke, G. & Struck, T. H. (2020). Delimitation of cryptic species drastically reduces the geographical ranges of marine interstitial ghost-worms (Stygocapitella; Annelida, Sedentaria). Molecular Phylogenetics and Evolution 143: 106663.
Struck, T. H. & Cerca, J. (2019). Cryptic species and their evolutionary significance. In: Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd: Chichester.
Cerca, J., Purschke, G. & Struck, T. H. (2018). Marine connectivity dynamics: Clarifying cosmopolitan distributions of marine interstitial invertebrates and the meiofauna paradox. Marine Biology 165: 123.
Struck, T. H., Koczula, J., Stateczny, D., Meyer, C. & Purschke, G. (2017). Two new species in the annelid genus Stygocapitella (Orbiniida, Parergodrilidae) with comments on their biogeography. ZooTaxa 4286: 301–332.
![]()