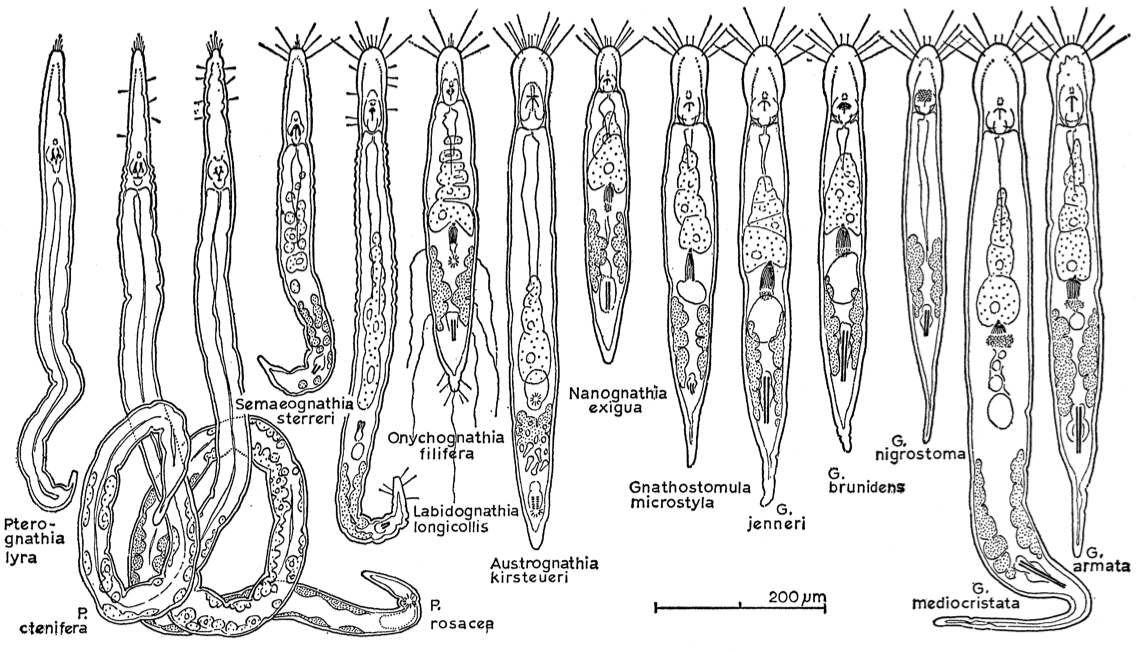
Gnathostomuida, also known as “lesser jaw worms”, is a phylum of about 100 described species of minute marine worms. The first gnathostomulid was discovered in 1928, in the fine sand of Kiel (Germany), by Adolf Remane. Later, Riedl (1969) elevated the animal group to the rank of phylum.
MINUTE WORMS WITH PRETTY INFORMATIVE JAWS
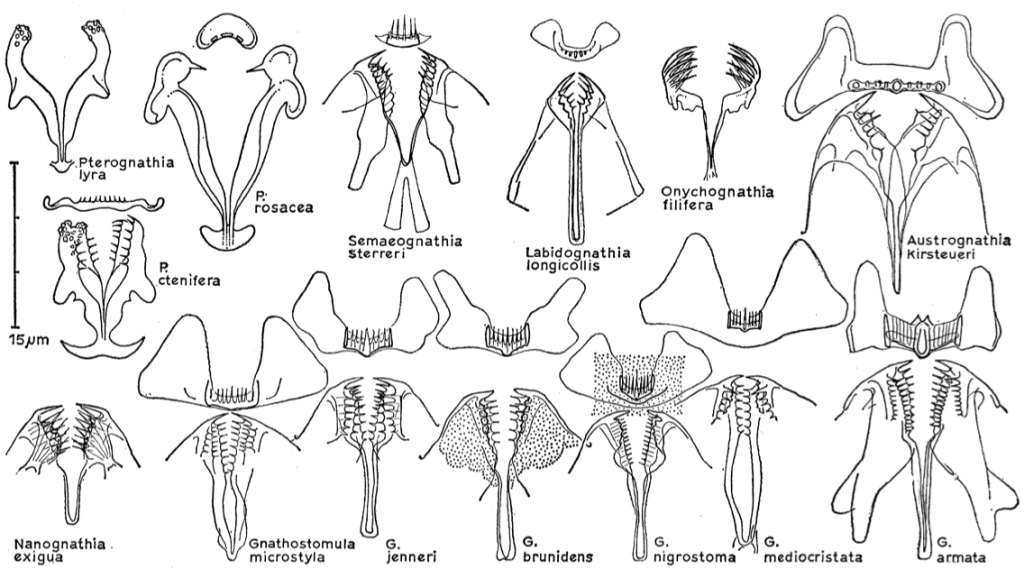
Even if quite small in terms of size, gnathostomulids exhibit a great morphological diversity in the foregut area (Riedl 1969). The mouth, located ventrally in the collar area, is hardened by cuticularized basal plates, sometimes with a “jugum” in the upper lip mostly with a basal plate. Thanks to the diversity of such jaw structures, the classification of the animals is possible. Presumably, this variety in the jaws shape may be an adaptation to different diets.
THE PHYLOGENOMIC PROBLEM
Originally ranked among Turbellaria, following studies keep arguing that there was a strong affinity between gnathostomulids and platyhelmints (Ax 1985, Meglitsch & Schram 1991, Peterson & Ernisse 2001). However, most of the molecular analysis of this time were based on a restricted taxon sampling that did not allow test of alternative hypotheses (Jenner 2004, Sterrer & Sorensen 2020). As an alternative hypothesis, Rieger (1976) signaled similarities in the ultrastructure of the monociliated epidermal cells in gastrotrichs and gnathostomulids. Some of the conflicting views that lie behind the similarities of the gnathostomulida group was summarized by Sterrer et al (1985), who concluded that gnathostomulids possesses traits that pointed towards platyhelmints, nemertodermatids, some groups of microscopic annelids, gastrotrichs, and rotifers (Sterrer & Sorensen 2020).
From the morphological point of view, such a dilemma was sort of solved by proposing a potential homology between the pharyngeal parts of gnathostomulids and rotifers (including acantocephalans). As a result, these two groups were united in a clade named Gnathifera (Alrichs 1995a). This putatively relationship obtained further support once the Micrognathozoa phylum was discovered in a number of studies based on morphological data (Haszprinar 1996, Kristensen & Funch 2000, Nielsen 2001, Halanych 2004).

Unfortunately, whereas the morphological approach appears to have reached a certain consensus about the phylogenetic position of gnathostomulids in the animal tree of life, molecular phylogenies still fail to present unambiguous results.
Some analyses of 18S rRNA suggest a sister-group relationship between gnathostomulids and chaetognaths (Littlewood et al. 1998, Glenner et al. 2004), but this result may well be a long-branch artifact (Jenner 2004). Other studies found an acoel-gnathostomulid relationship (Peterson & Eernisse 2001), and a monophyletic group consisting of Gnathifera and Cycloneuralia was recovered by Giribet et al. (2000, 2004). A later study from Dunn and coworkers recovered instead gnathostomulids in a clade with rotifers, acoels, and myzostomids (Dunn et al. 2008). In conclusion, a clear answer about which is the phylogenetic position of this obscure animal clade remains a mystery.
Having a solid solution to this phylogenetic issue, although it may first seem as uninteresting, may be extremely helpful while trying to better understand some key evolutionary questions such as how the ancestor of all Spiralia looked like and which was the lifestyle of it. This is one of the reasons why such an animal group is currently being study in our research group whiting the frame of the InvertOmics project.
If you want to know if we can bring some light to this interesting research question, please, keep yourself tuned!
BIBLIOGRAPHY
Ahlrichs, W. H. (1995a): Seison annulatus und Seison nebaliae. Ultrastruktur und Phylogenie. Ver. Deut. Zool. Gesell. 88: 155.
Ax, P. (1985): The position of the Gnathostomulida and Platyhelminthes in the phylogenetic system of the Bilateria. In: Morris, S,C., George, J. D., Gibson, R. & Platt, H. M. (eds.), The origins and relationships of lower invertebrates, pp. 168–180. The Systematics Association, Cambridge University Press, Cambridge.
Dunn, C. W., Hejnol, A., Matus, D. Q., Pang, K., Browne, W. E., Smith, S. A., Seaver, E., Rouse, G. W., Obst, M., Edgecombe, G. D., S.rensen, M. V., Haddock, S. H. D., Schmidt-Rhaesa, A., Okusu, A., Kristensen, R. M., Wheeler, W. C., Martindale, M. Q. & Giribet, G. (2008): Broad phylogenomic sampling improves resolution of the animal tree of life. Nature 452: 745–749.
Giribet, G., Distel, D., Polz, M., Sterrer, W. & Wheeler, W. C. (2000): Triploblastic relationships with emphasis on the position of Gnathostomulida, Cycliophora, Platyhelminthes and Chaetognatha; a combined approach of 18S rDNA sequences and morphology. Syst. Biol. 49: 539–562.
Giribet, G., Sorensen, M. V., Funch, P., Kristensen, R. M. & Sterrer, W. (2004): Investigations into the phylogenetic position of Micrognathozoa using four molecular loci. Cladistics 20: 1–13.
Glenner, H., Hansen, A. J., S.rensen, M. V., Ronquist, F. Huelsenbeck, J. P. & Willerslev, E. (2004): Bayesian inference of metazoan phylogeny: a combined molecular and morphological approach. Curr. Biol. 14: 1644–169 [erratum in Curr. Biol. 2005, Vol. 15, Issue 4].
Halanych, K. (2004): A new view of animal phylogeny. Annu. Rev. Ecol. Evol. Syst. 35: 229–256.
Haszprunar, G. (1996): Platyhelminthes and Platyhelminthomorpha – paraphyletic taxa. J. Zool. Syst. Evol. Res. 34: 41–48.
Jenner, R. A. (2004): Towards a phylogeny of the Metazoa: evaluating alternative phylogenetic positions of Platyhelminthes, Nemertea and Gnathostomulida, with a critical reappraisal of cladistic characters. Contrib. Zool. 73: 3–163.
Kristensen, R. M. & Funch, P. (2000): Micrognathozoa: a new class with complicated jaws like those of Rotifera and Gnathostomulida. J. Morphol. 246: 1–49.
Littlewood, D. T. J., Telford, M. J., Clough, K. A. & Rohde, K. (1998): Gnathostomulida – an enigmatic metazoan phylum from both morphological and molecular perspectives. Mol. Phylogenet. Evol. 9: 72–79.
Meglitsch, P. A. & Schram, F. R. (1991): Invertebrate zoology. Oxford University Press, Oxford.
Nielsen, C. (2001): Animal evolution. Interrelationships of the living phyla, 2nd edition. Oxford: Oxford University Press.
Peterson, K. J. & Eernisse, D. J. (2001): Animal phylogeny and the ancestry of bilaterians: inferences from morphology and 18S rDNA gene sequences. Evol. Dev. 3: 170–205.
Riedl, R. J. (1969): Gnathostomulida from America – this is the first record of the new phylum from North America. Science 163: 445–442.
Rieger, R. M. (1976): Monociliated epidermal cells in Gastrotricha: significance for concepts of early metazoan evolution. Z. Zool. Syst. Evol. 14: 198–226.
Sterrer, W., Mainitz, M. & Rieger, R. M. (1985): Gnathostomulida: enigmatic as ever. In: Morris, S. C., George, J. D., Gibson, R. & Platt, H. M. (eds.), The origins and relationships of lower invertebrates, pp. 183–199. Clarendon Press, Oxford.
Sterrer, W. & S.rensen, M. V. (2006): Chirognathia dracula gen. et spec. nov. (Gnathostomulida) from the west coast of N. America. Mar. Biol. Res. 2006: 296–302.

![]()