
Månedens gruppe jeg vil presentere for dere i dag, er en gruppe jeg har et nært personlig forhold til. Gruppen har en lang historie for meg. Jeg vil presentere familien til Annelida Dinophilidae.
Etter å ha skrevet diplomoppgave (i dag masteroppgave) om proteiner i et virus relatert til HIV ved Max Planck-instituttet for molekylær fysiologi i Dortmund, ønsket jeg å velge et annet tema for doktorgraden min. Jeg fikk aldri helt kontakt med temaet i diplomoppgaven min, og det fikk meg til å tenke over hva jeg brenner for innen biologi. Jeg innså at grunnen til at jeg ville studere biologi handlet om evolusjon og zoologisk systematikk. Dette fikk meg virkelig til å tenne på alle mine intellektuelle stempler. Derfor tok jeg kontakt med Prof. Westheide, leder for Zoological Systematic-gruppen ved universitetet i Osnabrück, og han foreslo ulike mulige doktorgradstemaer, blant annet en studie av meiofaunaen på Galapagosøyene. Det ene emnet som virkelig trigget meg, handlet imidlertid om progenetisk evolusjon av Dinophilidae og lignende taxa.
Hva er Dinophilidae?
Dinophilidae er såkalte interstitielle arter. Det betyr at de er så små at de kan bevege seg i rommet mellom sandkornene uten å flytte på dem. Kroppslengden varierer fra 1 til 3 mm, men hos Dimorphilus-artene er hannene så små som 50 µm. Disse Dimorphilus-dverghannerne er dermed de minste dyrene på jorden. De består egentlig bare av en sekk som er full av sædceller, med svært få nerveceller (ca. 50) og et kopulatorisk organ. Med organet sprøyter de sædcellene inn i kroppen til hunnen, som ofte er deres egen søster i den felles eggkokongen. Derfor er deres eneste formål i livet å levere sædceller og å dø etterpå.
Kroppsorganiseringen hos dinofilider, som ikke er dverghanner, er relativt lik, og de har alltid et hode etterfulgt av et segment med to ringer rundt munnen, deretter seks kroppssegmenter og den siste kroppsenheten med anus som kalles pygidium hos annelider. Munnens svelg er en massiv muskuløs tunge som har en unik organisering av muskelcellene. Hos dyr er fibrillene i muskelcellene vanligvis ansvarlige for bevegelsen av muskelcellen og er bare orientert i én retning. Fibrillene kan bare forkortes aktivt, men for å forlenge dem må en annen komponent (f.eks. en annen muskelcelle) strekke muskelcellen og dermed fibrillene i den. I denne svelgmuskeltungen er fibrillene orientert i to retninger vinkelrett på hverandre. Effekten er at disse muskelcellene ikke kan forkortes ved hjelp av fibrillene, fordi de to settene med fibriller motarbeider hverandre. Dette høres ut som en ulempe, men fører faktisk til at svelgmuskeltungen blir et svært stivt organ som gjør det mulig å skrape organisk materiale fra overflater.
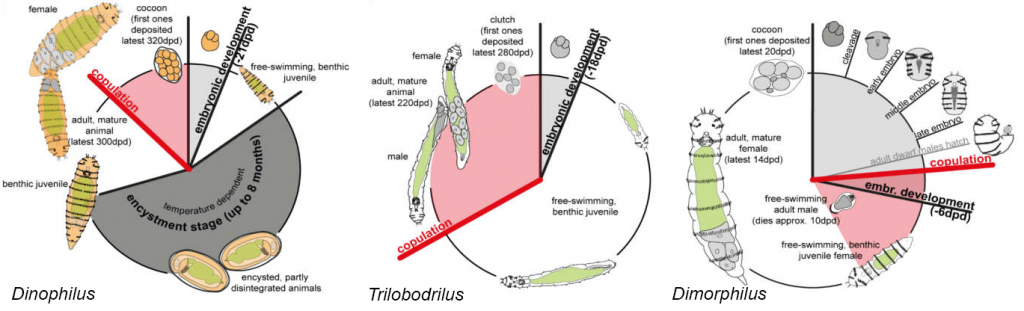
Familien består av tre slekter: Dinophilus med fem arter, Dimorphilus med to arter og Trilobodrilus med åtte arter. Livssyklusen til Dinophilus-artene tar omtrent ett år, noe som inkluderer tre til fire ukers embryonalutvikling og encystmentstadiet på opptil åtte måneder. På samme måte er livssyklusen til Trilobodrilus-artene også omtrent ett år, inkludert embryoutviklingen på to til fire uker. Et hvilestadium som hos Dinophilus mangler imidlertid. Livssyklusen til Dimorphilus er derimot mye raskere, med en livssyklus på rundt tre uker for hunnene, inkludert seks dagers embryoutvikling. Livssyklusen til dverghannerne er enda raskere med omtrent én uke.
Hva er progenetisk evolusjon?
Gould introduserte progenetisk evolusjon (eller progenese) som en av de evolusjonære prosessene som oppstår ved endringer i den relative utviklingshastigheten til de ikke-reproduktive versus de reproduktive kroppsdelene (eller mer vitenskapelig den somatiske versus den reproduktive utviklingen). Progenese er en av de to prosessene som fører til pedomorfose, noe som betyr at kroppen til etterkommeren ligner mer på kroppen til forfedrenes unge eller larvestadium enn den voksne kroppen til forfedrene. I progenese skjer dette ved at utviklingen av forplantningsorganene akselererer, mens utviklingshastigheten til de ikke-reproduktive organene forblir uendret. Derfor blir slike arter kjønnsmodne i ungdoms- eller larvestadiet og stopper sin videre utvikling. De ser ut som juvenile stadier eller larvestadier og har også en lignende kroppsstørrelse som disse stadiene. Den andre prosessen som fører til pedomorfose, er derimot neoteni. Ved neoteni bremses utviklingen av de ikke-reproduktive kroppsdelene (eller deler av dem), mens den reproduktive utviklingen skjer i normal hastighet. Slike arter ser også ut som forfedrenes juvenile stadier eller trekk, men har kroppsstørrelsen til det voksne stadiet. Mennesker har angivelig slike neoteniske trekk. Et eksempel er menneskets evne til å leke selv i voksen alder.

Hvordan kommer progenetisk evolusjon og dinofilider sammen?
På grunn av sin enkle kroppsbygning ble dinofilidene først ansett for å være flatormer, men så ble de anerkjent som ringormer. På grunn av den enkle organiseringen ble de imidlertid igjen ansett for å være et eksempel på ringormenes forfedre, noe som gjenspeiler den evolusjonære utviklingen fra enkel til kompleks, slik man ofte antar i dyrenes evolusjon. Og som så ofte før var denne antakelsen feil. Detaljerte morfologiske studier kunne vise at de faktisk er sekundært forenklede organismer, som sannsynligvis har utviklet seg ved progenese for å muliggjøre en raskere tilpasning til det interstitielle riket.
På grunn av deres likhet med larvestadiene i Eunicida-ordenen, som blant annet omfatter
som inkluderer tilstedeværelsen av to ringer rundt munnen, ble det foreslått
at de tilhører denne ordenen, nærmere bestemt familien Dorvilleidae.
Dette er svært interessant, ettersom denne ordenen også omfatter noen av de
største ringormene på jorden, for eksempel sandstrimmelormen (se en video av dem her; vidoen er på engelsk), som kan bli opptil 6 m lange. Den nøyaktige plasseringen av Dinophilidae i familien Dorvilleidae var imidlertid usikker.
Meg selv, dinofilider og progenetisk evolusjon – en lang historie
Det er her historien min begynner. Tanken med doktorgradsavhandlingen min var å bruke molekylære data for å vise hvor de er plassert i Dorvilleidae, og basert på denne plasseringen undersøke prosessen med deres progenetiske evolusjon nærmere. Men som så ofte i livet går det ikke som planlagt. I løpet av studiene ble det mer og mer åpenbart at Dinophilidae sannsynligvis ikke er en del av Eunicida i det hele tatt. Derfor var jeg nødt til å begynne å se på hele livets tre av annelider og ikke bare deler av det. På slutten av doktorgradsarbeidet mitt kunne jeg bare vise at de ikke er eunicider, men jeg kunne ikke plassere dem mer nøyaktig i annelidenes livstre med sikkerhet. I avhandlingen min nevnte jeg, med hensyn til de morfologiske likhetene, at de også kunne finnes i en annen annelidefamilie, Orbiniidae.
Ettersom jeg i løpet av doktorgradsstudiet utvidet mitt arbeid til å omfatte hele livstreet til annelidene, fortsatte jeg å jobbe med dette de neste årene etter doktorgraden, i tillegg til plasseringen av annelidene i dyrenes livstre. Til dette brukte jeg mer og mer data og sofistikerte metoder, og etter hvert tok jeg i bruk fylogenomiske metoder. Det betyr at jeg samlet inn data på genomskala for å besvare spørsmålene mine. På den tiden betydde dette ikke hele genomet, men transkriptomet. Ettersom dette fortsatt var ganske utfordrende for så små skapninger, måtte jeg utvikle nye metoder for å gjøre dette for andre mikroskopiske dyregrupper. Etter at jeg klarte å bruke disse metodene på så små organismer, gikk jeg tilbake til temaet for doktorgradsavhandlingen min. Og vi lyktes; vi kunne endelig plassere Dinophilidae i annelidenes livstre. Det var interessant at det viste seg at de hadde utviklet seg ved progenetisk evolusjon fra en orbiniid-lignende forfader. Dermed sluttet på en måte sirkelen for min reise med dinophilider siden jeg avsluttet doktorgraden min mer enn ti år tidligere. I studien kunne vi også vise at noen få andre interstitielle annelidgrupper også utviklet seg ved progenese, mens andre utviklet seg ved trinnvis miniatyrisering. Det var altså to evolusjonære prosesser som drev tilpasningen til det interstitielle området.
Der trodde jeg at spørsmålet jeg forsøkte å besvare i doktorgraden min endelig var avklart etter mer enn et tiår, men så kom Martin-Duran og hans medforfattere i 2020. Ved å publisere det første genomet til en dinofilid, analyserte de også plasseringen av Dinophilidae og de andre interstitielle gruppene på nytt. Mens de bekreftet vår tidligere posisjon for de fleste av de interstitielle gruppene, ble Dinophilidae plassert annerledes. Sammen med en annen interstitiell familie, Lobatocerebridae, ble Dinophilidae plassert som søster til størstedelen av det biologiske mangfoldet av annelider, en gruppe som kalles Pleistoannelida og som også omfatter både Orbiniidae og Eunicida. I dette scenariet har de sannsynligvis fortsatt utviklet seg ved progenese, men gitt denne plasseringen kan vi ikke fastslå det morfologiske utseendet til denne forfedren slik vi kan med de to andre plasseringene; morfologisk til Eunicida før og i våre analyser til Orbiniidae. Selv 25 år etter at jeg startet min doktorgrad i år, gjenstår fortsatt spørsmålet om hvor Dinophilidae skal plasseres i annelidenes livstre.
Det andre spørsmålet er derfor: Kommer jeg til å pensjonere meg på grunn av dette spørsmålet, som for alvor startet min akademiske karriere?
Tre morsomme fakta å huske på:
- Dinophilide dverghanner er de minste dyrene på jorden og lever bare for å ha sex.
- Dinofilider er egentlig larvestadier av en annelideforfader som ble kjønnsmoden. Det verste tilfellet av for tidlig tenåring man kan tenke seg.
- Dinofilider har fibriller som står vinkelrett på hverandre i den samme muskelcellen i svelgbollen. Dette gjør bulben til et svært effektivt skrapeorgan.
Bakgrunnslitteratur om dinofilider (på engelsk):
- Martín-Durán, J.M., B.C. Vellutini, F. Marlétaz, V. Cetrangolo, N. Cvetesic, D. Thiel, S. Henriet, X. Grau-Bové, A.M. Carrillo-Baltodano, W. Gu, A. Kerbl, Y. Marquez, N. Bekkouche, D. Chourrout, J.L. Gómez-Skarmeta, M. Irimia, B. Lenhard, K. Worsaae, and A. Hejnol, Conservative route to genome compaction in a miniature annelid. Nature Ecology & Evolution, 2020. https://doi.org/10.1038/s41559-020-01327-6
- Purschke, G., Anatomy and ultrastructure of ventral pharyngeal organs and their phylogenetic importance in Polychaeta (Annelida) – I. The pharynx of the Dinophilidae. Zoomorphology, 1985. 105: p. 223-239. https://doi.org/10.1007/BF00311966
- Struck, T.H., A. Golombek, A. Weigert, F. A. Franke, W. Westheide, G. Purschke, C. Bleidorn, and K.M. Halanych, The evolution of annelids reveals two adaptive routes to the interstitial realm. Current Biology, 2015. 25(15): p. 1993-1999. http://dx.doi.org/10.1016/j.cub.2015.06.007
- Struck, T.H., K.M. Halanych, and G. Purschke, Dinophilidae (Annelida) is most likely not a progenetic Eunicida: Evidence from 18S and 28S rDNA. Molecular Phylogenetics and Evolution, 2005. 37: p. 619–623. https://doi.org/10.1016/j.ympev.2005.07.010
- Worsaae, K., A. Kerbl, M.D. Domenico, B.C. Gonzalez, N. Bekkouche, and A. Martínez, Interstitial Annelida. Diversity, 2021. 13(2): p. 77. https://www.mdpi.com/1424-2818/13/2/77
- Westheide, W., Progenesis as a principle in meiofauna evolution. Journal of Natural History, 1987. 21: p. 843-854. https://dx.doi.org/10.1080/00222938700770501
Teksten er oversatt fra den engelske versjonen ved hjelp av DeepL.
![]()