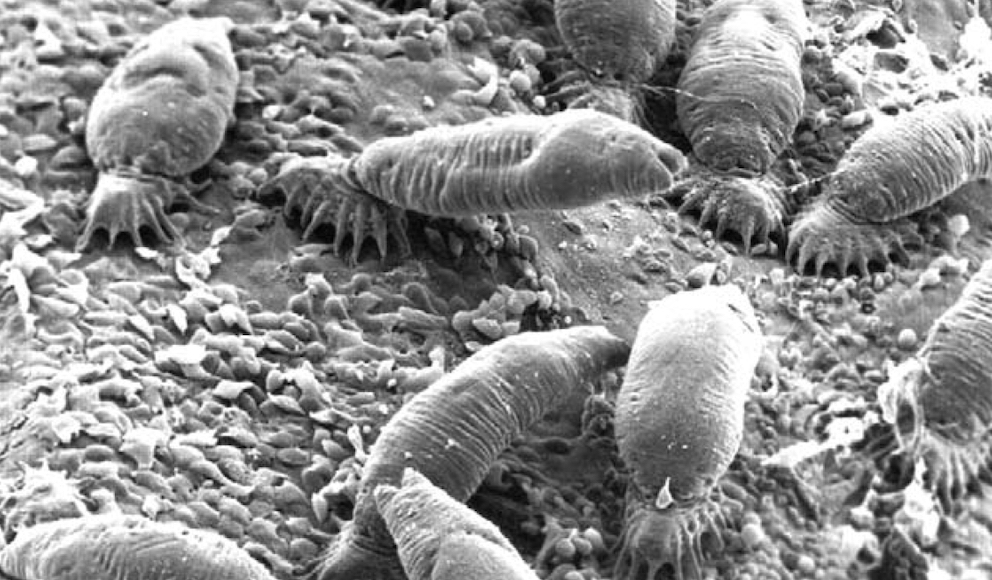
Monogeneans are a group of parasitic flatworms (Platyheminthes). The group is species-rich and systematists assign them to a class within the flatworms. Together with flukes (class Trematoda) and tapeworms (class Cestoda) they form the superclass Neodermata. The name-giving feature Neodermata is that the original epidermis is replaced by a new neodermis. Monogenean flatworms are ectoparasites that occur on the skin or gills of fish, and some species may cause substantial harm to their hosts. A well known example is Gyrodactylus salaris on Atlantic salmon (Salmo salar); in particular in Norway G. salaris has been a threat to wild salmon populations. However, except for those species that cause substantial harm to hosts that are of economic importance Monogenea usually do not attract much attention, not even among biologists. Accordingly, the community of scientists doing research on Monogenea is also pretty small.
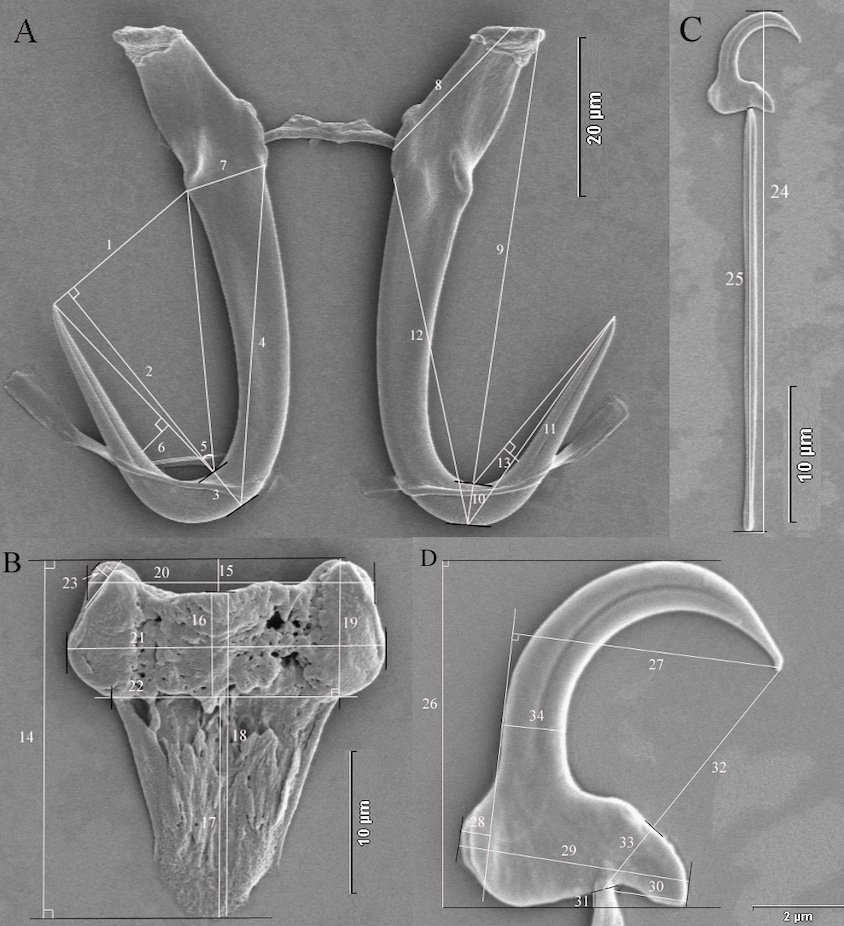
Monogenea are small and at first glance pretty unimposing animals, even the ‘larger’ species are usually below 1 cm in length. As compared to the Trematoda and Cestoda the monogenean flatworms have the simplest parasitic life cycle. They develop directly and, usually, do not change the host; in fact, many species are host specific. Infection of new host individuals is mostly by direct (skin) contact. The most prominent morphological features of monogenean worms are sclerotized structures at the back end of the animals that allow the parasites to stay attached to the host – biologists refer to it as an opisthaptor. This organ consists of various sickles, hooks, hamuli and other attachment structures. Figure 1 illustrates these structures for a particular strain of G. salaris in large magnification. Taxonomists are particularly interested in these structures, as they turned out very useful diagnostic features for species description and identification. Many species are only found on one particular host species (specialists), while others have been observed on various host species (generalists). There is also increasing evidence that parasites may explore new host species and subsequently adapt to them (host switching).
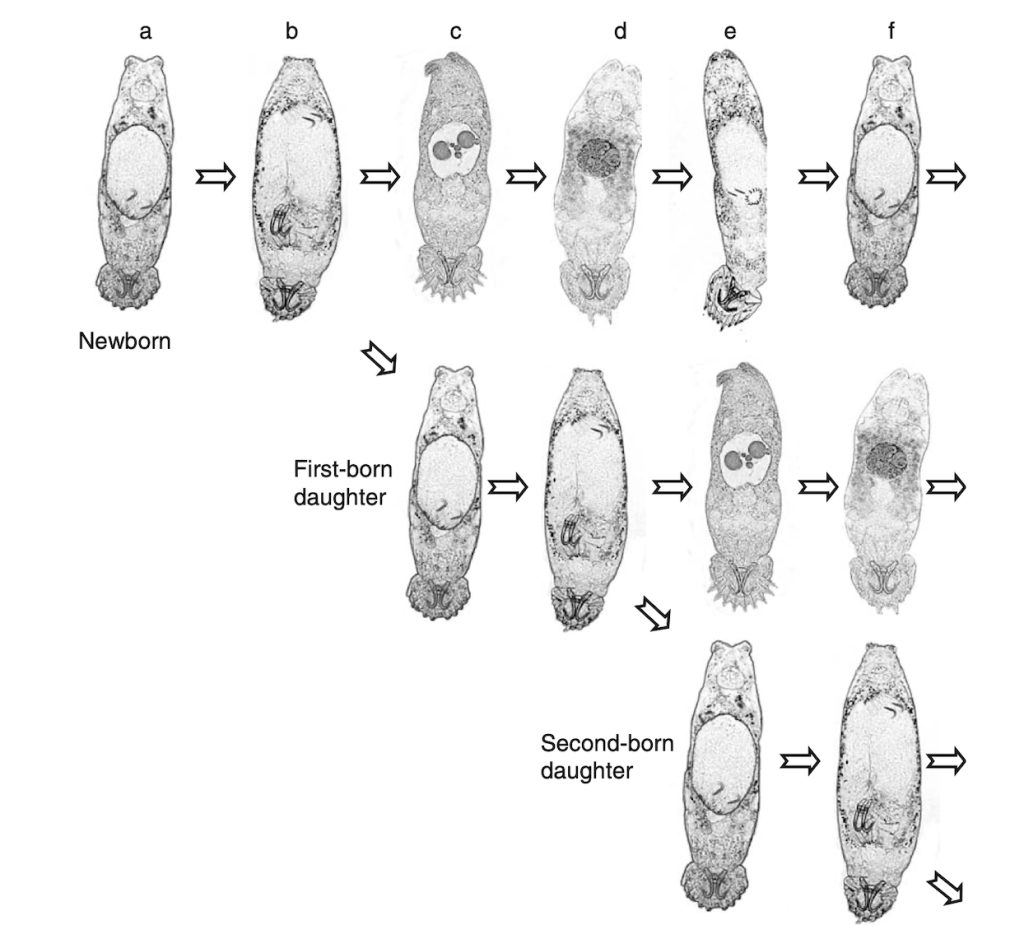
While some monogenean species are ‘egg-laying’ and release the eggs directly into the surrounding water, there is many viviparous species with the new born larvae immediately attaching to the host. The species of the genus Gyrodactylus are viviparous, and the above mentioned Gyrodactylus salaris became also known as the ‘Russian-Doll killer’. As explained in Figure 2, a ‘newborn’ parasite will soon give birth to a new parasite (first-borne daughter) before a new individual develops and so on. The infection of the host can, thus, reach soon a high number of parasites. However, most species infect their hosts in only low to moderate numbers.
A very interesting question is the biodiversity of monogenean flatworms. Currently, scientists have no reliable estimates of how many monogenean flatworm species there are. In the literature one finds estimates of several thousands of species, but all scientists agree that the majority of monogenean species are not yet discovered and described. Traditionally, the hard parts of the opisthaptor and the host species are most important for species identification, but recently molecular data were increasingly used (see also DNA barcoding). These DNA sequence data indicated for several genera that the number of species is, indeed, underestimated. For example, most fish species that were screened for the presence of Gyrodactylus species were infected by at least one species, frequently specific for the host fish. As an interesting intellectual game one may extrapolate the currently available information on Gyrodactylus species diversity and conclude that one may expect several ten-thousands of Gyrodactylus species, while hitherto only several hundreds have been described. If indeed true, Gyrodactylus species may turn into model organisms for evolutionary studies and processes that lead to the formation of different species.
Research on parasitic flatworms has a long tradition at the Natural History Museum Oslo (NHM), and also FEZ member Lutz Bachmann has conducted several projects. Interestingly, the genome of Gyrodactylus salaris has been deciphered at NHM (Hahn et al. 2014), which paved the way for many downstream discoveries. A future FEZ project on flatworms will address the impact of so-called mobile genetic elements, i.e. DNA that can change position within a genome, on the evolution of monogenean flatworms.
References:
Bakke TA, Cable J, Harris PD. (2007). The biology of gyrodactylid monogeneans: the “Russian-doll killers”. Advances in Parasitology 64:161-376. https://doi.org/10.1016/S0065-308X(06)64003-7
Hahn C, Fromm B, Bachmann L. (2014) Comparative genomics of flatworms (platyhelminthes) reveals shared genomic features of ecto- and endoparastic neodermata. Genome Biology and Evolution 6: 1105-1117. https://doi.org/10.1093/gbe/evu078
![]()